|
|
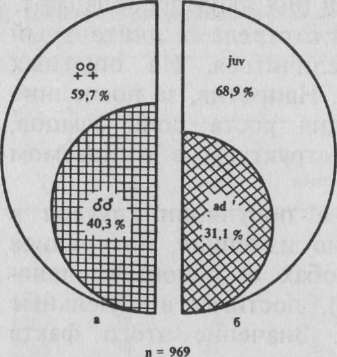
Главная → Справочник → Статьи → Блог → ФорумСтруктура популяцийПод экологической структурой популяции обычно понимают определенное соотношение возрастных групп и полов, сочетание оседлых животных с животными-мигрантами, наличие семейных и прочих группировок. Экологическая структура в значительной мере определяет жизнеспособность популяции и отражает процессы, происходящие в популяции: успешность размножения, степень подвижности, динамику численности и условия существования, в целом (Шварц, 1969). Непременными характеристиками любой популяции живых организмов являются особенности полового и возрастного состава и связанная с ними динамика репродукции поголовья. При всяком популяционном анализе эти характеристики следует учитывать (Тимофеев-Ресовский и др., 1973). При исследовании отдельной популяции крайне необходимо учитывать структуру и следить за ее изменениями. К сожалению, в отношении глухаря характеристики структурных особенностей поголовья в литературе либо не представлены, либо даны с большими оговорками, что не позволяет рассчитывать на надежность приводимых сведений. Изучение животных в состоянии их естественной свободы требует применения определенных приемов охоты или способов отлова, чтобы иметь пробы из популяции и по ним судить о структуре поголовья. В идеальных пробах соотношение тех или иных групп в выборке должно быть таким же, как и в природе. С. С. Шварц указывает, что промысел всегда приводит к изменению половой и возрастной структуры популяции, поскольку большинство способов и приемов охоты выборочно. На самом деле так случается далеко Не всегда. Изучая соотношение полов у глухаря, Ю. Н. Киселёв (1971) сопоставил данные осенних учетов на ленточных пробах с материалами осеннего отлова живоловушками за 6 лет и пришел к заключению, что эти материалы абсолютно одинаково характеризуют изменения соотношения полов. В исследованиях, проводимых в Кировской области при отлове живых глухарей, кривые уловистости птиц (живо-ловными устройствами) и возрастного состава популяции (процент молодняка) вполне синхронны. Что же касается полового состава, то здесь данные, если они представлены десятками особей, тоже отражают действительное соотношение полов в природе, но с некоторыми поправками. В осенний период самцы не так прочно привязаны к определенным участкам территории, как весной, а после окончания линьки и распада выводков более равномерно распределены по угодьям. Однако анализ повторных встреч показал, что самцы чаще, чем самки, попадаются в ловушки. Каждый самец был повторно пойман в среднем 1,8 раза, самка 1,4 раза, при максимальном количестве поимок соответственно 8 и 4. Можно предположить, что самцы менее осторожны, чем самки, охотнее идут в ловушки, а поэтому чаще попадаются в стационарные самоловы. На самом деле выборочность при отлове глухарей универсальными живоловушками может несколько исказить действительную картину соотношения полов в природе, завысив количество взрослых самцов из-за их меньшей подвижности. Применение однотипных орудий лова год за годом в одних и тех же местах даже при наличии некоторых погрешностей выборки должно отражать основные тенденции и характер изменений структуры популяции за ряд лет, а для разных районов — специфические ее черты, поскольку степень отклонения полученных данных от истинных остается неизменной. Проводя отлов глухарей осенью шторными ловушками, Ю. Н. Киселев (1971) установил, что половая структура популяции глухаря в Окском заповеднике изменяется в больших пределах. Ежегодно среди пойманных птиц там преобладали то самцы, то самки, хотя в некоторые годы (1963 и 1965) последние составляли подавляющее большинство. Отлов птиц этими же ловушками весной приводит иногда к заметному преобладанию самцов в пробе. В условиях Кировской области весной (в мае) в ловушки попадаются (за редким исключением) только самки. Анализируя анкетные сведения по учету тетеревиных птиц в Волжско-Камском регионе за 1965 г. (730 анкет), Е. Г. Киселева и С. Г. Приклонский (1971) обнаружили, что относительное число самцов уменьшается к южной границе ареала с 58,7 до 46,2% и наиболее резко в Ульяновской области —до 38,6%. В районах преобладания самцов плотность населения, как правило, выше: на 10-километро-вом маршруте отмечено в среднем 3,1 птицы, там же, где преобладали глухарки, — в среднем 2,8 (разница статистически недостоверна). Данные, полученные в Кировской области (одной из самых северных областей Волжско-Камского региона), правда, на небольшом материале (36 особей), не согласуются с приведенными выше: именно в 1965 г. в отлове было отмечено максимальное преобладание самок (75%). Противоречат этим сведениям и материалы Окского заповедника (Рязанская область), свидетельствующие о том, что в южной части ареала самцы, как правило, преобладают в популяции, а выход молодняка сравнительно низок (Киселев, 1971). В Печоро-Илычском заповеднике (Коми АССР), по данным 5405 встреч за 1937—1939 гг., 58% составляли самцы и 42 —самки (Теплов, 1947). Однако в выводках из 111 особей самок было 55%. Автор, признавая, что самок рождается больше, объясняет это большей частотой их гибели от хищников, несмотря на покровительственную окраску. В районе заповедника Денежкин Камень в 1954 г. самцы при учете по встречам составляли 56%, в б. Висимском заповеднике в 1948 г.— 50%, а в открытом для охоты районе г. Полевского в 1949-1953 гг.— 32% (Данилов, 1963). Автор считает, что в последнем случае нормальное соотношение полов было нарушено в результате охоты. По данным анализа крыльев глухарей в 1967—1970 гг. (Финляндия), соотношение полов среди молодых глухарей 3 осенних сезона было равным, и в одном сезоне преобладали самки; среди старых птиц самки . преобладали 2 сезона, и 2 сезона соотношение полов было равным. По данным учета в 1969 г., среди взрослых глухарей самки составляли 61,4% и среди молодых — 56,2%, в 1970 и 1971 гг. соответственно 61,6 и 57,7%. Осенью 1952— 1961 гг., по результатам анализа 6734 крыльев глухарей, собранных от охотников, определено, что в добыче преобладали самцы. Среди взрослых птиц их было 56%, среди молодых — 58%. Расхождения материалов по учету и анализу крупных промысловых проб объясняются выборочностью отстрела. В условиях стационара (Кировская область) в осенний период охотники также отстреливают больше самцов, преимущественно молодых, а весной на токах —только взрослых. Однако в течение ряда лет охоту на глухаря весной и осенью практически не проводили, а при отлове живых гггиц живоловушками в популяции по-прежнему продолжали преобладать самки. Генетический механизм обеспечивает расщепление потомства по полу в соотношении 1:1 (Тимофеев-Ресовский и др., 1973). Указанное первичное соотношение вследствие различной выживаемости мужских и женских особей может измениться при вылуплении из яиц (вторичное соотношение), а также в течение жизни взрослых половозрелых особей. Соотношение полов у глухаря при рождении пока не изучено. До того момента, когда можно определить пол у этой птицы, в опытах С. П. Кирпичева (личное сообщение) в 4 кладках выжило 18 самок и 12 самцов, т. е. наметилось преобладание самок в выводках, ранее отмеченное для верховьев р. Печора (Теплов, 1947). Немногочисленные наблюдения на опытных участках Кировской области при встречах взматеревших выводков в августе, когда удается визуально определить пол птенцов, также свидетельствуют, что самки в них явно преобладают. Вследствие выборочности отлова и отстрела за длительный срок процент самок должен увеличиться. На опытных участках этого пока не произошло. Напротив, за последние годы наблюдается явная тенденция роста доли самцов, а не самок. Изменений половой структуры в ожидаемом направлении не наблюдается. Показатель возрастного состава популяции глухаря в Кировской области также довольно изменчив, но среднее количество молодых особей в пробах из района исследований составляет почти % (66,5%), достигая в отдельные годы 82% от общего поголовья. Значение этого факта состоит прежде всего в том, что он указывает на высокие воспроизводственные способности популяции и в конечном итоге характеризует условия обитания. С другой стороны, высокий процент молодняка в сентябре и октябре свидетельствует о хорошей его сохранности в течение всего выводкового периода, а также в начале осени. На одну взрослую самку в Кировской области приходится в среднем 3,6 молодых. Характерны сравнительно небольшие колебания возрастного состава по годам. В отдельные годы отклонения процента молодых особей в популяции от средней многолетней составляют максимум 24,4% (1976 г.), обычно же не превышают 10—12%. Благодаря этому обеспечивается ежегодно почти стабильный уровень воспроизводства популяции с увеличением общего количества особей к осени в 3 раза. Преобладание самок (56,6%) оказывает положительное влияние на уровень воспроизводства в условиях полигамии. Анализируя приведенные данные, следует указать, что материал не всегда отвечает требованиям статистики. В частности, данные за 1975 и 1983 гг. нельзя рассматривать при характеристике структуры поголовья данного года, но для анализа в целом за весь период они пригодны. Осенние материалы о половом и возрастном составе поголовья глухарей в Кировской области показали, что в структуре популяции произошли заметные перемены. Самцы стали в отлове преобладать над самками (52,6%), в то время как средний многолетний (за 10 лет) процент их равен 40,3%- Доля самок упала соответственно с 59,7 до 47,4% (рис. 1).
Рис. 1. Половой (а) и возрастной (б) состав популяции глухаря в бассейне р. Вятка в среднем за 11 лет
Количество молодняка к осени 1976 г. сократилось весьма существенно — до 42,1%, а за 6 лет —до 62,5% (за период 1964—1974 гг. этот показатель колебался от 80,0 до 51,4%, в среднем составляя 68,9%). К осени 1980 г. прирост численности оказался наибольшим (81,8%) за все годы наблюдений. В составе пробы снова преобладали самцы —53,1%. В хозяйстве областного общества охотников и рыболовов среди 30 глухарей, отловленных осенью 1980 г., было 4 взрослых и 10 молодых самцов, 11 взрослых и 5 молодых самок. Процент самцов составил 46,7%, молодых и взрослых птиц оказалось поровну. На изменение полового и возрастного состава популяции глухаря отрицательное влияние могут оказывать чрезвычайно неблагоприятные погодные условия. Так, например, в весенне-летний период 1976 г. наблюдалась массовая гибель глухарей. Однако катастрофического снижения численности, которого следовало ожидать из-за плохих погодных условий, все-таки не произошло. Вероятно, общая гибель молодняка и части взрослых птиц не превысила 50% поголовья. Были основания надеяться, что в благоприятных условиях численность популяции может восстановиться в течение одного репродуктивного цикла. Осенью 1977 г. структура популяции глухаря начала выправляться. Доля молодых увеличилась почти в полтора раза —с 42,1 до 62,9%, а количество молодых птиц, приходящихся на одну взрослую самку, —с 1,6 до 2,9. Однако ни один из этих показателей не достиг средней многолетней величины. Рост численности глухаря в 1977 г. происходил на фоне обильного урожая брусники и рябины, молодняк рос и развивался ускоренно. К началу октября отдельные молодые самцы достигли веса 3450 г (хотя обычно их вес в это время не превышал 3300 г). Данные, полученные в результате определения пола и возраста глухарей по крыльям от 11 птиц, добытых в научно-опытном охотничьем хозяйстве и в других районах об ласти (от 15 птиц), были сопоставлены с материалами по отлову живых глухарей. В результате выяснилось, что в отлове за 1977 г. самцы составляли 48,1%, в ружейной добыче —58%, а молодняк в пробах соответственно 62,9 и 61 5%. Расхождения в большей степени касаются половой структуры, в то время как показатели возрастного состава из проб ружейной и самоловной добычи оказались весьма близкими. По погодным условиям весь бесснежный период 1978 г. во многом повторил экстремальные условия 1976 г. Как и тогда, весна была затяжной и холодной, такое же холодное было и лето. Чрезмерное обилие осадков, вызвавшее неоднократный подъем паводковых вод, ранние заморозки и снег, выпавший 1 октября, — таковы характерные приметы репродуктивного сезона. Все это не могло не вызвать снижения показателей интенсивности размножения глухарей, поскольку условия инкубации, питания птенцов и взрослых птиц в весенне-летний период были далеки от оптимальных. Лютая стужа зимой 1978—1979 гг. с морозами до 48—50°С, резкое потепление и чрезмерно полноводный весенний разлив в начале мая при относительно благоприятных условиях в период инкубации также оказали отрицательное влияние на результаты размножения, снизив долю молодняка от обычного уровня на 10%. В эти годы наблюдалось незначительное увеличение доли самцов (в пределах 10—12%) по сравнению со средним многолетним их значением. Показатели отлова живых глухарей на путиках и анализ половозрастной структуры популяции свидетельствовали, что высокая численность и плотность населения птиц наблюдались к осени 1980 г., а в последующем году произошел некоторый спад. Общее некоторое снижение численности произошло в результате уменьшения доли молодняка, обусловленное характером погодных условий. В отличие от предыдущего года репродуктивный период 1981 г. отличался запоздалой и очень дружной весной, высоким паводком и чрезвычайно засушливым летом. Короткая весна в сочетании с летней засухой снова вызвали снижение доли молодых особей в популяции почти на 1/3— с 76,4 до 58,8%. Сопоставление материалов, полученных путем отлова глухарей живоловуш-ками и отстрела по годам, показало, что различия между данными этих двух способов получения проб не столь существенны, поэтому в представительности тех и других не приходится сомневаться. Аналогичное явление отмечено В. Г. Телепневым в Западной Сибири, хотя и в меньшей степени. Там популяция молодых глухарей сократилась с 58,5 до 48,6%. Несмотря на сокращение доли молодых, численность глухаря осенью 1981 г. как в Кировской области, так и в Западной Сибири находилась на достаточно высоком уровне благодаря сохранности маточного поголовья после благоприятного для размножения года. Отлов живых глухарей на научно-опытном участке указывает на очень высокую долю взрослых самцов, что можно объяснить результатом направленного вмешательства (в последние годы ни один взрослый глухарь не был вывезен за пределы хозяйства), а также тем, что в 1980 г. в пробе из популяции было необычно много молодых самцов (35,2%), которые взматерели, но сохраняются до сих пор. К осени 1982 г. прирост популяции (по данным отлова) оказался весьма низким (48,5%). Независимо от степени достоверности приведенных материалов можно предположить, что в 1982 г. наблюдались спад численности глухаря и изменение структуры популяции в сторону заметного увеличения доли взрослых самцов. По данным В. Г. Телепнева, структура населения глухаря в подзоне средней тайги Западной Сибири (Томская область) характеризуется преобладанием самок, но рост численности к осени обеспечивается в среднем менее чем в 2 раза. В бассейне р. Нарым на 1 взрослую самку приходится в среднем 1,2 птенца. В подзоне южной тайги Западной Сибири (Новосибирская область) за 14 сезонов на маршруте (с использованием снегохода) общей протяженностью 15 тыс. км было встречено 662 глухаря, в том числе 498 самцов (75,2%) и 164 самки (24,8%), причем среди отстрелянных 116 птиц самцы и самки находились в таком же соотношении. Громадное преобладание самцов в северном районе Барабинской степи, В. Г. Телепнев объясняет отсутствием охоты и усиленным влиянием хищников (соболь, ястреб-тетеревятник, филин), которые успешно справляются с самками глухаря (копалухами). На юге ареала, в недостаточно благоприятных условиях (отсутствие камешков-гастролитов в грунте, ограниченные площади сосняков), подобное состояние популяции соответствует оптимальной плотности населения глухаря на невысоком уровне. Ю. Н. Киселев на материалах отлова глухарей в течение 6 лет показал, что в Окском заповеднике, на южном пределе распространения птиц в европейской части страны, также наблюдается преобладание самцов при сравнительно низком (43,6%) ежегодном приросте поголовья. В Северо-восточных районах Финляндии, по данным, приведенным Р. Л. Потаповым (1985), состав популяции глухаря в пробе, полученной с помощью отстрела, характеризуется невысоким процентом молодых особей —54,3 при такой же доле самок. В Финляндии относительное количество молодых птиц в популяции глухаря (за 7 лет) изменялось от 65 до 39%, в СССР на р. Печора (за 6 лет) —от 80 до 30%, Лапландском заповеднике, СССР (за 14 лет) —от 81 до 7%, в Окском заповеднике (за 6 лет) — от 67 до 14%. В отличие от соседних популяций, в вятской тайге не наблюдается ни резких скачков численности, ни падения интенсивности размножения, что придает динамике численности более плавный характер (рис. 14). В бассейне р. Вятка в настоящее время популяция глухаря находится в благоприятных условиях. Такие же условия существуют в отдельных районах Среднего Урала, где выход молодняка достаточно высок и численность глухаря к началу осенней охоты иногда увеличивается на 280%. Известно, что молодые птицы обладают пониженной воспроизводственной способностью и плодовитостью. У глухаря это, очевидно, является главным фактором, определяющим интенсивность размножения. Степень участия молодых самцов и самок в размножении в известной мере предопределяет результаты репродуктивного цикла. Молодые особи в возрасте 11 мес в данном случае составляют своеобразный резерв популяции, который в зависимости от плотности населения и условий окружающей среды может участвовать в размножении в большей или меньшей степени. «Холостые» глухари, которых часто отмечали в районе р. Печора и значительно реже на Среднем Урале,— это именно молодые птицы, не участвовавшие в размножении.
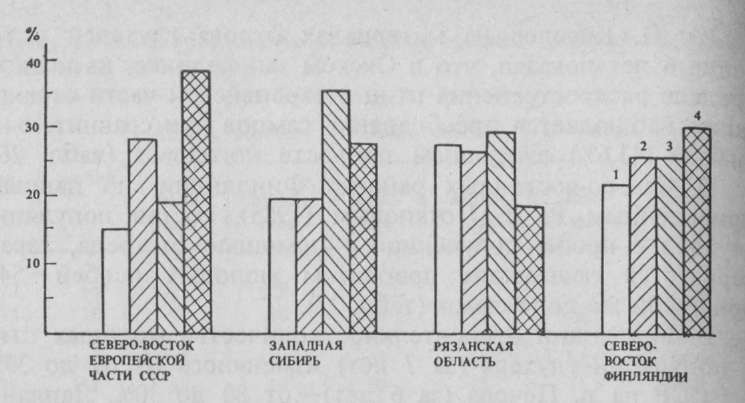
Рис. 2. Доля взрослых (/) и молодых (2) самцов, взрослых (J) и молодых (4) самок в популяциях глухаря из различных частей ареала
Молодые (11-месячные) самцы способны к размножению, но не принимают активного участия в токовании и, как правило, не «поют» при наличии поблизости взрослых доминантных самцов. 18 мая был на току молодой петушок, который «запел» после того, как были отстреляны все взрослые самцы, посещавшие ток. В последние годы стало известно 3 случая добычи глухарей на токах в возрасте 1 года. Это служит сигналом о недостатке (перепромысле) взрослых самцов на токах. Самки глухарей способны к спариванию и яйцекладке в возрасте 1 года, однако гнездовой инстинкт у них несколько ослаблен и отчетливо проявляется лишь во вторую весну их жизни. В опытах В. В. Немцева и др. в питомнике глухарки приступали к размножению не раньше второй весны, но в возрасте 2 лет участвовали в размножении только 57% птиц. В естественных условиях молодые самки иногда приступают к размножению, но позднее взрослых. Взрослые самки с ярко выраженным инстинктом насиживания физиологически лучше и раньше подготовлены к яйцекладке. Они активнее молодых на току, поэтому быстрее выбирают партнера для спаривания, в лучшие сроки завершают кладку и насиживание и успешнее выращивают птенцов. Очевидно, большая часть брошенных кладок принадлежит также молодым самкам, которые иногда отказываются от насиживания. Преобладание самок в составе популяции глухаря в Кировской области не является патологическим явлением. Отчасти поэтому популяция обладает повышенной репродуктивной способностью и высокими темпами размножения. Характеристики полового и возрастного составов вместе взятые показывают, что в условиях подзоны южной тайги глухарь может производить значительное количество биомассы в виде ценной охотничьей продукции. Графический анализ структуры популяции по годам показал, что существует прямая зависимость между величиной годового прироста и количеством молодых самцов (рис. 2). Иными словами, в годы подъема численности глухаря среди молодняка, как правило, больше самцов, а в годы, когда наблюдается некоторое снижение интенсивности размножения — процент молодых самцов осенью низкий. Подобное явление отмечено в Томской области, где молодые особи среди самцов составляли 21,7%, среди самок — 80,2%. Пока неизвестно, получается ли это вследствие неодинаковой рождаемости или повышенной естественной смертности одного из полов. Тем не менее следует признать, что повышенная рождаемость самок или высокая смертность самцов в первые 3 мес постэмбрионального развития в конечном итоге приводят к одинаковым результатам: увеличению уровня плодовитости на следующий год. По мнению автора, это является важным (но не единственным) свойством популяции как саморегулирующейся системы. Изучение изменений структуры популяций глухаря следует проводить в более широких масштабах. Особенно перспективны исследования в периферийных частях ареала где численность птиц изменяется резко и в больших пределах, а регулирующие механизмы проявляются более четко и их легче раскрыть. Сочетание оседлых и мигрирующих особей как один из признаков, относящихся к структуре популяции, заслуживает отдельного рассмотрения, поскольку глухарь в этом плане почти не исследован. Границы и размер популяций. Н. В. Тимофеев-Ресовский, А. В. Яблоков, Н. В. Глотов справедливо отмечают, что сведений о размерах популяций глухаря накоплено крайне мало, но полагают, что установить границы популяций ориентировочно можно с помощью наблюдений за перемещениями большого числа меченых особей в течение всей их жизни. Попыток установить границы отдельных популяций глухаря никто не предпринимал, несмотря на то что современный ареал обыкновенного глухаря, особенно в южной и западной его частях, в значительной степени расчленен (см. рис. 1). В условиях изоляции отдельных лесных массивов, очевидно, могли сформироваться популяции со специфическими особенностями, но по фенотипи-ческим признакам эти популяции не всегда можно выделить, а их эколого-физиологические характеристики в сравнительном аспекте слабо изучены. Анализируя факты самых дальних перелетов глухарей, переместившихся далее 8 км и добытых далеко за пределами участка кольцевания, нельзя не отметить, что все перелеты имели направление на запад, северо-запад или восток. Ни одна из птиц не направилась на юг, где на протяжении десятков километров глухарей практически нет. Причина этого кроется не только в различии угодий, но и в существовании четко выраженного пространственно-механического барьера, который глухари, как правило, не преодолевают. Таким барьером служит в данном случае долина р. Чепца, луговая пойма которой имеет местами ширину более 2 км. За поймой по левому берегу проходит железнодорожная магистраль, за которой по пойменной террасе простираются пахотные земли с отдельными куртинами лесов, сохранившихся по понижениям вдоль ручьев, балкам и оврагам. Сельскохозяйственные угодья левобережья р. Чепца имают более половины всей площади, а елово-пихтовые 33.р0Вные леса с примесью березы прерывисты и не образуют массивов, площадь которых превышала бы 3-5 км2. Б угодьях левобережья р. Чепца обычен тетерев, очень редок рябчик и отсутствует глухарь. Отдельные птицы иногда все же проникают в эти угодья, но закрепиться гам они не могут. В. А. Агафонов сообщил, что он наблюдал самца глухаря в стае тетеревов на убранном зерновом поле. Южнее в аналогичных стациях известны случаи отстрела двух одиночных глухарей (самца и самки), но сведений о постоянном местообитании и глухариных токах отсюда не поступало. Эту обширную территорию глухари не заселяют естественным путем на протяжении десятков лет. Изредка проникающие одиночные особи-мигранты в конце концов исчезают бесследно или бывают отстреляны. В северной таежной части научно-опытного охотничьего хозяйства по правобережью р. Чепца лесные массивы разобщены в меньшей степени, но и здесь обширные поля и вырубки вокруг д. Роговая создают некоторое препятствие для расселения глухарей. Через безлесные пространства, как показывают наблюдения, глухари не летят, а огибают их с двух сторон. Чаще всего птицы, окольцованные на опытном участке, улетают на запад, в угодья соседнего приписного охотничьего хозяйства «Меховщик». Здесь они распространяются по полосе сосновых лесов вдоль боровой террасы р. Чепца, минуя неширокие полевые и луговые угодья в нижнем течении р. Светлица. Из одного лесного массива в другой птицы проникают там, где смыкаются хвойные леса. Грунтовые дороги, неширокие реки и лесные покосы не являются преградами для расселения глухарей. Таким образом, в районе научно-опытного хозяйства реально существует одна граница популяции — южная. Во все остальные стороны происходит свободное взаимное проникновение и перемешивание особей из различных токовых группировок. Этим обеспечивается поддержание численности на определенном уровне. Указанная граница создалась в результате распашки земель. В современных условиях пространственно-механи-ческие барьеры для глухаря создаются в районах сплошных концентрированных вырубок, приводящих к образованию временных пустошей. Они разобщают популяции и нарушают сложившиеся ранее связи на длительный срок. Вопрос о величине отдельных популяций глухаря, которые могут существовать в условиях пространственной изоляции, чрезвычайно интересен, но и труден. Если рассматривать популяцию как «элементарную размножающуюся единицу вида», то территорию, тяготеющую к определенному току, можно рассматривать как арену жизни элементарной популяции, но, как правило, леса площадью 1000 га и даже более, в которых имеются все необходимые условия для жизни глухарей, эти птицы не заселяют. Очевидно, при искусственном расселении глухарей, помимо характера самих угодий, следует учитывать и величину лесного массива. Минимальная площадь абсолютно пригодных угодий, на которых возможно длительное существование изолированной популяции глухаря, не должна быть меньше 4000 га. На такой площади возможно существование 4 отдельных, но общающихся между собой токовых группировок. Высказанные соображения были учтены при обследовании места выпуска глухарей в двух районах Литовской ССР, которые при однотипности условий различались величиной лесного массива — 3000 и 12 000 га. Предпочтение было отдано последнему (пуща Шимонис). На территории исследования в Кировской области имеется один ток, который существует в неполном окружении пахотных земель. Он расположен на болоте около р. Роговка, максимальная ширина его составляет 2 км. С лесным массивом ток связан перемычками леса вдоль речки вверх и вниз по ее течению. С востока ток и основной лес разъединены полосой пахотных земель сложной конфигурации, но нигде не превышающей по ширине 1 км. Как выяснилось, глухари иногда перемещаются к лесу и обратно. Чаще всего глухари проникают в большой массив леса и обратно через его выступы. Очевидно, длительное существование этого глухариного тока в небольшом массиве, да еще поблизости (3 км) от крупного села в известной степени обусловлено тем, что возможности перемешивания особей здесь не нарушены. Границы и размеры популяции глухаря в условиях тайги очертить пока невозможно. Вместе с тем можно с уверенностью сказать, что отдельные самостоятельные (островные) популяции глухаря в пределах его ареала сформировались в результате изменений среды обитания преимущественно под влиянием антропогенных факторов. Сравнительно небольшие группировки птиц, образовавшиеся после дробления лесных массивов (из-за освоения новых земель, мелиорации угодий и по другим причинам), лишились ряда преимуществ. Они потеряли широкие связи, а вместе с ними и широкую экологическую пластичность вследствие обеднения наследственной основы. Можно предположить, что это служит одной из причин такого, казалось бы, парадоксального явления, когда численность глухаря, например, в Центральной Европе, несмотря на многолетнее запрещение охоты, не увеличивалась. Обыкновенного глухаря в центральных районах европейской части СССР характеризуют двумя основными признаками: преобладанием самок и высокими темпами размножения (увеличение численности в 3 раза). В периферийных районах ареала обыкновенного глухаря, наоборот, преобладающее значение в популяциях приобретают самцы, а уровень воспроизводства обеспечивает увеличение численности к осени только в 2 раза. Половой и возрастной состав популяций глухаря в Кировской области представляется оптимальным. В нем заложены и реализуются значительные резервы для восполнения поголовья. Усилия, предпринятые для того, чтобы найти и очертить сколько-нибудь заметные границы отдельных популяций внутри естественного ареала глухаря, также не увенчались успехом. Во всех остальных регионах воспроизводственные показатели снижены вследствие менее благоприятных условий обитания, вызванных производственной деятельностью человека. КомментарииСтатьи по теме:
Главная → Справочник → Статьи → Блог → Форум |
|
|
|
© Все права защищены.
Копирование материалов невозможно. |
|